
期刊:
臨床研究期刊 (The Journal of Clinical Investigation) Volume 117 Number 5 May 2007
標題:
以中和抗體抑制腫瘤成長因子β 可避免因放射治療引起的腫瘤加速轉移
(Inhibition of TGF-β with neutralizing antibodies prevents radiation-induced acceleration of metastatic cancer progression)
作者
Swati Biswas,1 Marta Guix,2 Cammie Rinehart,2 Teresa C. Dugger,2 Anna Chytil,1
Harold L. Moses,1,3,4 Michael L. Freeman,5 and Carlos L. Arteaga1,2,4
單位
屬於美國田納西州 那什維爾 范德堡大學(Vanderbilt University)
1.癌症生物學系、2.醫學系、3.病理學系、4. Vanderbilt-Ingram癌症中心乳癌研究計畫 以及 5.放射腫瘤學系
摘要
我們探究了因癌症治療而產生的TGF-β究竟會不會加速腫瘤的發生。我們使用了MMTV/PyVmT 此一轉移型乳癌基因的轉殖模型,指出了游離輻射(譯註1)或doxorubicin(譯註2)都會增加循環中TGF-β1與腫瘤細胞的量,並會增加肺部的癌症轉移。這些效應都因為我們使用了泛TGF-β抗體而受抵銷。我們在有TGF-β抗體的環境下,體外培養了這些在循環中會表現多瘤病毒中等T抗原(譯註3)的腫瘤細胞,發現它們不會生長;這表示TGF-β在這些細胞中是一種生存的訊號。當老鼠的腫瘤不含有第二型TGF-β受器時,放射線就不會增加肺部的癌症轉移;這顯示至少在部分上,肺癌轉移是跟TGF-β在腫瘤細胞上的直接作用有關的。這些數據含示,癌症治療所產生的TGF-β是一種幫助癌症轉移的訊號,也提供了一個原理根據,讓我們得在癌症治療時同時使用TGF-β抑制劑。
譯註1:此為放射性治療。
譯註2:此為一癌症化學治療常用藥物,俗稱「小紅莓」。
譯註3:此為感染此病毒之初期的產生的三種T抗原中其中一種。
介紹
腫瘤成長因子TGF-β,既是一個腫瘤抑制基因,也是一個腫瘤啟動子。當TGF-β配體(ligands)結合到與其對應的絲胺酸/酥胺酸(Ser/Thr)激酶跨胞膜受器後,這些受器接著就會磷酸化、活化Smad族的訊息傳遞者。被活化之後,Smad 2、Smad 3 會與 Smad 4 結合並遷移至細胞核內,在此處調控與細胞週期停止(cell cycle arrest)、細胞凋亡(apoptosis)相關的基因之轉錄。TGF-β之所以有腫瘤抑制基因之用,即係此項調控之功。的確,在內皮細胞與基質中的TGF-β訊息傳遞,若是喪失或減弱,都會讓內皮細胞的細胞惡性轉化(transformation)大行其道。另一方面,已有研究顯示,若將顯性且無效的TGF-β受器引入至移轉的癌症細胞中,從上皮細胞型態轉變成間葉細胞型態的轉分化(epithelial-to-mesenchymal transdifferentiation)、能動性(motility)、侵略性(invasiveness)、以及存活率(survival)均會受到抑制。這證明在已經完全轉化的細胞中,TGF-β扮演了一個腫瘤啟動子的角色(於參考文獻4中有回顧)。大部分的癌都保有TGF-β受器,卻喪失或部分喪失了Smad依賴性的抗細胞分裂效應(Smad-dependent antimitogenic effect);同時,在某些個案中,他們還會對TGF-β有所反應,而獲得了幫助癌症轉移的能力。進而,不論癌症細胞過度生產TGF-β、過度活化TGF-β,或是兩者兼備,都會藉由旁泌(paracrine)的方式,改變腫瘤周圍的微小環境(microenvironment),幫助腫瘤的發生。這些資料提供了支持根據,讓我們得以設法阻斷人類癌症中的自泌性(autocrine)或旁泌性(paracrine)之TGF-β訊息傳遞,以達治療之目的。
除Smad族的蛋白質外,TGF-β還可以刺激數種轉化的訊息傳遞路徑。之前TGF-β已被證實可保護轉化細胞(transformed cells)不受細胞凋亡之害。此類細胞反應有一種可能的機制,就是TGF-β誘發了PI3K和PI3K的標的物,也就是絲胺酸/酥胺酸激酶 Akt,此訊息傳遞程序和對於癌症藥物的抗藥性是相關連的。有些對傳統化學療法有抗藥性的腫瘤會過度表現TGF-β,而TGF-β的抑制劑經證明可逆轉此種抗藥性。還有,大部分的腫瘤都會過度表現TGF-β配體(ligands)。在腫瘤組織與在血清中的高量TGF-β配體,與腫瘤轉移的復發、患者較差的癒後,均有所相關。
在乳癌的基因轉殖模型中,TGF-β之訊息傳遞可加強轉移既存的乳腺腫瘤。這是受例如Neu/ErbB2或多腺瘤病毒中央T抗原(polyomavirus middle T antigen, PyVmT)等的癌症基因之誘發所致。再者,在MMTV/LTR乳腺啟動子的控制下,表現PyVmT癌症基因的基因轉殖鼠,經過有條件的TGF-β1誘導,只要兩個禮拜之短,轉移肺癌的機率便高了十倍。有些癌症治療已被證實會全面性地、或留在原處誘發TGF-β。所以我們推測,被癌症治療所誘發的TGF-β,會提供存活訊息給這些對化療有抗藥性的腫瘤裡的細胞,或者一部分此種腫瘤中的細胞,使它們可能在治療之後,立即加速腫瘤的擴展。在此,我們使用了乳癌轉移的MMTV/PyVmT 基因轉殖模型來展示,游離輻射(ionizing radiation)和小紅莓(doxorubicin)提高了循環中的TGF-β1濃度、循環中的腫瘤細胞數、以及肺部的癌症轉移。這些效應都藉由TGF-β中和抗體的施加,而得以抵銷。在缺乏第二型TGF-β受器(TβRII)的老鼠身上,放射線則沒有增加肺部的癌症轉移。這些資料暗示,癌症治療所產生的TGF-β是腫瘤中一個幫助癌症轉移的訊號,因此也提供了原理根據,使我們在使用這些療法時,得以同時搭配使用TGF-β的抑制劑。
結果
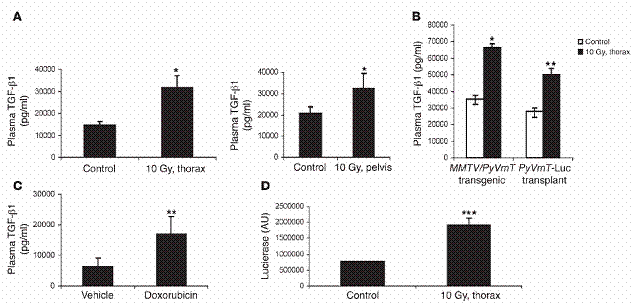
胸腔放射線處理與化學治療提高了循環中的TGF-β量。我們在八週大的未交配母鼠之胸腔與骨盆內,施加了10 Gy的放射線。在施加放射線之後隔24小時,進行血液採集。我們在受放射處理之老鼠的血漿裡,觀察到約為控制組2倍的TGF-β1含量 (胸腔,P=0.03;骨盆,P=0.02;圖一A);同時,TGF-β2的量沒有改變(資料未呈現)。在八週大的MMTV/PyVmT基因轉殖鼠中,或是在種下MMTV/PyVmT腫瘤細胞、穩定地轉染了luciferase 表現載體的非基因轉殖鼠中,我們都得到類似的結果(分別為P=0.015與P=0.007,相較於控制組,圖一B)。在經過放射處理後,TGF-β1的濃度有持續七天比控制組高(資料未呈現)。為了將這些結果推廣到其他的癌症治療法,我們測試了DNA嵌入劑(DNA-intercalating agent)與topoisomerase II之抑制劑 doxorubicin(Adriamycin)的效果如何。從第八週開始,基因轉殖鼠被施行三次腹腔注射doxorubicin (5 mg/kg i.p.),每次分別間隔21天。在第十五週收集到的血漿裡發現,相較於未處理的老鼠,TGF-β1濃度提高了兩倍(P = 0.009;圖一C),而TGF-β2濃度則是持平不變。為了要測量在放射處理後五週所取得的肺臟組織中,被活化的TGF-β1之濃度,我們使用了一種TGF-β1生物檢驗來檢測,這檢測是使用一種會穩定表現plasminogen activator inhibitor-1/luciferase reporter (PAI-1/luciferase reporter) 的貂肺內皮細胞來運作的。相較於未受放射處理之老鼠肺組織裂解液,經過放射線處理的老鼠肺組織裂解液,誘導提升了有2倍多的TGF-β1濃度(P=0.0008;圖一D)。
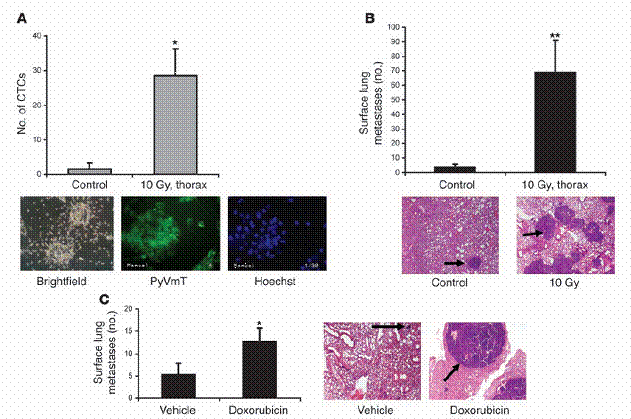
胸腔的放射線處理及化學治療,會增加循環中的腫瘤細胞量與增加肺部癌症轉移。TGF-β增強了會表現PyVmT腫瘤基因之腫瘤細胞的存活率,而且之前已被證實,TGF-β有條件地被誘發後,會在MMTV/PyVmT基因轉殖鼠中加速乳腺腫瘤的轉移。因此,我們推測,因治療而提升的循環中TGF-β1濃度,可能和這個乳癌轉移模型中的循環中腫瘤細胞增加、肺部癌症轉移增加,有所關連。帶有腫瘤的MMTV/PyVmT老鼠在第八週大時,被施行胸腔放射處理(thoracic irradiation)(10 Gy)直至第十三週為止。實驗終結時進行心臟穿刺以取得血液,而血液中的細胞部分,則拿去測試其在活體外生成細胞集落之能力。從未經放射處理之老鼠所取得之血液,產生了平均1.6 ± 1.5個細胞集落;而從經過放射處理之老鼠所取得之血液,於10-12天後所測量的結果是,產生了平均28.3 ± 7.6個細胞集落(P=0.02:圖二A)。PyVmT抗體的免疫染色證實,這些生長的細胞集落有表現癌症基因(圖二A)。循環中的活腫瘤細胞數有所增加;而和此相符合的是,我們在經放射處理的老鼠中,觀察到較未經處理之控制組17倍的肺部表面癌症轉移(P = 0.009;圖二B)。在被用doxorubicin處理相較於未處理之老鼠中,也在肺部癌症轉移觀察到類似的上升情形(P = 0.032;圖二C)。在施加doxorubicin 24小時之後,相較於未經處理之控制組老鼠所取得之血液,也觀察到在細胞群落數目的上升(17.2 ± 3.7與7.2 ± 2 colonies; n = 5; P = 0.007)。從老鼠的體重及一系列的腫瘤直徑數據來看,受處理與未處理的老鼠在腫瘤負荷量上並無差異,這表示TGF-β1濃度之差別,不能簡單地以腫瘤負荷量會隨時間增加來解釋。
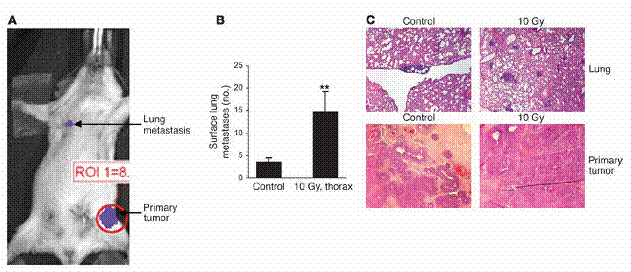
接下來,我們檢驗了放射線所誘發之肺部癌症轉移,是否僅限於帶有腫瘤的MMTV/PyVmT基因轉殖鼠身上。為了測試這個可能性,我們從會表現PyVmT之基因轉殖的乳腺癌中,取得表現luciferase的腫瘤細胞,然後將其注射至非基因轉殖的同源FVB老鼠身上。這些老鼠中的腫瘤,生長在左側鼠蹊(inguinal)(四號)的乳腺脂肪墊,我們以生物發光法,觀察並估計了其轉移擴散至肺臟的情形(圖三A)。當腫瘤長到了200mm3或更大時,老鼠在胸腔施加了10 Gy 的放射線並在其餘部分施加屏障,兩週後將老鼠犧牲。相較於控制組,接受放射處理的老鼠增加了三倍的表面肺部癌症轉移(P = 0.005; 圖三,B和C)。再者,在放射處理後24小時與兩週之後,分別有相較於控制組多8倍與5倍的循環中腫瘤細胞量(表一)。在實驗組與控制組間,原發腫瘤的生長率並無不同(資料未呈現)。
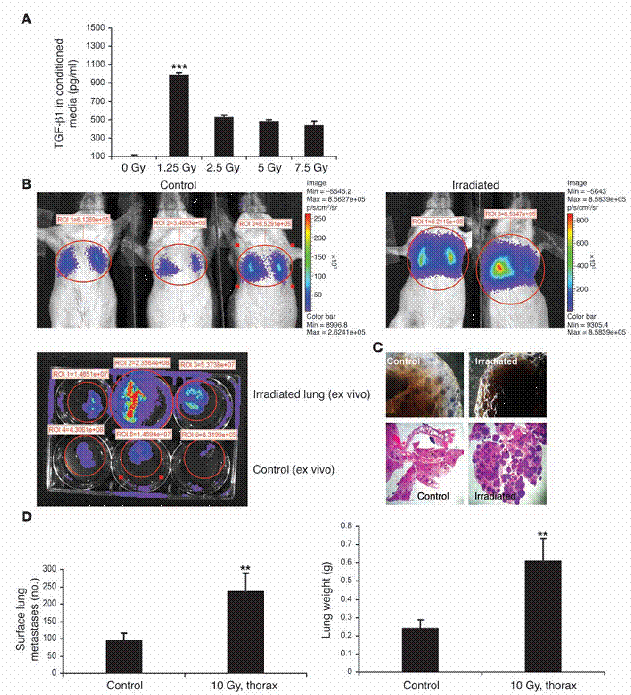
將培養中的PyVmT細胞暴露在1.25 - 7.50 Gy下,會增加其產生TGF-β的量(圖四A)。在以上所述之研究過程中,有既成之腫瘤的老鼠才會施加放射線處理;循環中TGF-β濃度的增加,有可能是因為放射線在癌症細胞上的作用,因此我們想排除掉這個可能。我們對未交配的FVB母鼠施加了10 Gy的胸腔放射。在放射處理後一個小時,我們從尾部靜脈注入了表現 PyVmT 且受 luciferase 穩定轉染(stably transfected)的腫瘤細胞。肺臟中的腫瘤細胞負荷量(tumor cell burden),是在腫瘤細胞的注入之後,使用活體內(in vivo)與活體外(ex vivo)的生物發光來監測的。經放射線處理的老鼠肺臟,表現了較控制組為高的生物發光訊號(圖四B),此訊號與人工計數之表面肺癌轉移數量是相關連的,亦與肺臟重量相關(圖四,C與D)。
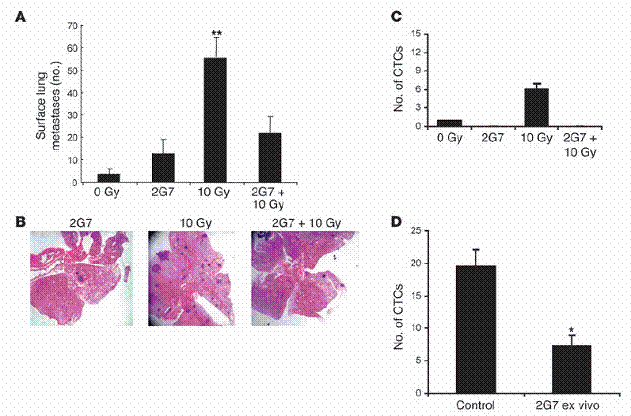
TGF-β中和抗體阻斷了受放射線誘導增加之肺部癌症轉移。我們使用了2G7中和泛TGF-β IgG2抗體 (2G7 neutralizing pan-TGF-β IgG2)來驗證,放射線所誘導增加的循環中TGF-β濃度,是否在肺癌轉移中扮演一個成因的角色。這個單株抗體阻斷了所有三種哺乳類所具有的TGF-β異構物,並且在活體中有活性。八週大的MMTV/PyVmT基因轉殖鼠,在胸腔放射處理的2小時之前,以15mg/kg 之2G7或PBS注射至腹腔中。這些處理持續每週兩次,到第十三週為止,此時將老鼠犧牲並測量是否肺部有癌症轉移。然而有趣的是,施打2G7到未經放射線處理的老鼠中,少許增加了肺癌轉移,但在統計上並不顯著(圖五A和B)。在兩種實驗組別之間,並未存在原發腫瘤細胞負荷量的差異(資料未呈現)。
我們收集了血液並體外培養其細胞懸浮液,以估計在循環中的腫瘤細胞量。從未經放射處理與經過放射處理的老鼠身上,血液中取得而培養的細胞群落數,均因施加2G7而顯著地減少(圖五C)。最後,我們檢驗了在體外添加2G7以抑制TGF-β,是否會阻止老鼠血液中取得之腫瘤細胞群落生長。將帶有腫瘤、經過放射處理的老鼠所取得之血液,接種於含有2G7與不含有2G7的環境下,而在十天之後評估其集落生長情形。將細胞培養在存有2G7的環境下,發現會降低70%的細胞群落數(圖五D),這表示自泌性的(autocrine) TGF-β對於表現PyVmT的循環中癌症細胞,是一種存活因子。
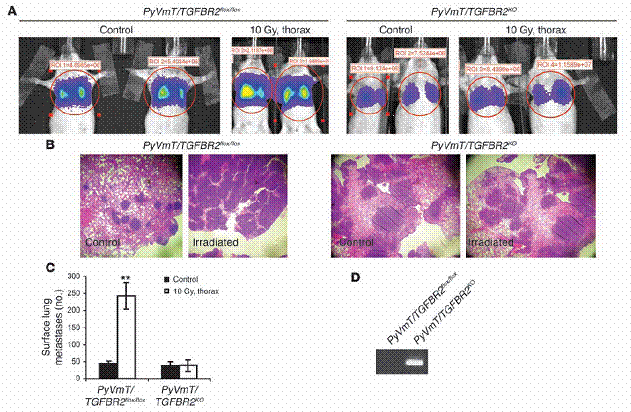
放射線誘導增加肺部癌症轉移,需要腫瘤細胞內的TGF-β受器。以上描述的結果顯示,因放射線誘導之循環中TGF-β濃度提升,是對於循環中腫瘤細胞的存活信號,讓細胞可以移居至肺臟、在肺臟裡生長。因此我們推論,藉由移除細胞對循環中TGF-β的反應,以使得TβRII 不存在於MMTV/PyVmT腫瘤中,就可以抵銷放射處理在癌症轉移上的效應。為檢驗此推論之可能,我們使用了表現PyVmT的腫瘤細胞,並且用Cre-Lox技術將TβRII有條件地於細胞中移除。含有或未含TGFBR2的PyVmT表現型腫瘤細胞株(在此之後將稱為PyVmT/TGFBR2flox/flox or PyVmT/TGFBR2KO)是分別從PyVmT/TGFBR2flox/flox 及 PyVmT/TGFBR2KO老鼠身上的乳腺癌所產生。PyVmT/TGFBR2flox/flox 及PyVmT/TGFBR2KO細胞株均以luciferase之載體穩定轉染,且經由尾部靜脈注射入八週大、未交配的FVB母鼠,這些母鼠有些沒有經過放射線處理,有些則是在注射腫瘤細胞之前1小時,施加了10 Gy的胸腔放射。兩週後估計肺部癌症轉移情形。在被注射PyVmT/TGFBR2flox/flox 細胞的老鼠裡,經放射處理之老鼠相較於控制組老鼠,發現生物發光訊號有所增加、肺部癌症轉移增加6倍;但如果老鼠之前是被注射不含有TβRII的細胞,在施加與未施加放射處理的兩組中,便沒有差別(圖六,A-C)。PyVmT/TGFBR2KO 的腫瘤結節比 PyVmT/TGFBR2flox/flox 的腫瘤結節還大,此結果與之前的一項報告是相符合的,也就是說,PyVmT表現型乳癌細胞之TβRII喪失,會增進肺部癌症轉移的生長。經PCR擴增腫瘤組織中取得之DNA證實了在 PyVmT/TGFBR2KO細胞中,是存在基因重組的(圖六D)。
討論
我們在基因轉殖老鼠的乳癌轉移模型中,研究了癌症治療所誘發的TGF-β是否會加速腫瘤的發生。在MMTV/PyVmT基因轉殖鼠中,在肺部施加游離輻射或是以doxorubicin處理,都增加了循環中的TGF-β1量、循環中腫瘤細胞數以及肺部癌症轉移。循環中TGF-β1濃度增加、之後的癌症轉移增加,並不需要老鼠在施加處理時就擁有腫瘤,就如同我們也在放射處理後才施打腫瘤細胞的老鼠中所觀察到的。TGF-β1會增加,並不僅限於受到胸腔放射處理的誘導,因為我們在骨盆的放射處理、以及全面性施加doxorubicin下,也都觀察到了TGF-β1的受誘導增加。在放射處理後,循環中腫瘤細胞的增加以及腫瘤轉移的增加,都因到2G7的施加而抵銷。重要的是,循環中PyVmT表現型腫瘤細胞無法在有2G7的環境下進行體外培養,這也就表示了自泌性(autocrine)TGF-β對這些細胞是一種存活的訊號。這些結果與之前兩項研究是相吻合的;在這些研究中,以可溶性TβRII:Fc融合蛋白、或用反義TGF-β1(antisense TGF-β1)穩定性轉染加以阻斷TGF-β,都抑制了腫瘤細胞的能動性、存活率、滲入(intravasation)以及肺部癌症轉移。
老鼠身上的腫瘤不含TβRII,放射線便無法在這些老鼠身上加強肺部癌症轉移。為了展示此點,我們從PyVmT/TGFBR2flox/flox 與 PyVmT/TGFBR2KO之老鼠身上既成之乳腺腫瘤,取得細胞株來使用。Forrester等人曾經提出報告,以MMTV/Cre表現有條件地使TGFBR2缺失時,這些knockout PyVmT表現型的腫瘤發生潛伏期就變短了許多,並且表現出明顯增加的肺部癌症轉移,這是相較於PyVmT/TGFBR2flox/flox型的腫瘤。因為我們使用了靜脈注射法,並且,我們所注入細胞株是從既成之含有TβRII與未含TβRII的腫瘤而來,所以我們的結果無法解釋Forrester等人所報導之癌症潛伏與轉移的表現。儘管我們在經放射處理的老鼠中無法排除TGF-β之增加,是否會影響宿主的微環境(microenvironment)與/或免疫系統,並進而幫助了腫瘤轉移之進行,我們不含有TβRII之腫瘤的結果卻強烈地顯示,癌症轉移之增加至少在部分上,是肇因於TGF-β在癌症細胞的直接作用。
在腫瘤細胞中,這些潛在受循環中TGF-β濃度調控的訊號反應,目前仍屬未明。然而,TGF-β的助癌轉移效應並不僅限於此基因轉殖模型。舉例說明,在MMTV/LTR啟動子控制下表現有活性的TGF-β1與Neu的雙基因型(bigenic)的老鼠,也表現出比MMTV/Neu更多的循環中腫瘤細胞與肺部癌症轉移。會表現Neu和具活性的TGF-β1之乳腺腫瘤細胞,能侵入Matrigel並且能在transwells上移動,但卻受可溶性的TβRII:Fc給阻斷,表示單單Neu不足以誘導出一種侵略性的表現型。再者,在會表現具活性Neu的基因轉殖腫瘤裡,若共同表現具活性的突變種Alk5和TβRI,會增加肺部血管外的癌症移轉(extravascular lung metastasis),這與TGF-β之效應相符:也就是腫瘤周圍蛋白酶、癌症細胞的附著和侵略。PyVmT是一個強而有力的癌症基因,我們也已經知道它會活化ErK和PI3K;但2G7竟然能對如此的腫瘤細胞具備有效的抗腫瘤移轉效果,有點不符合直覺。但是,這表示TGF-β可以放大癌症基因的訊號,而只要這訊號大於某一閾值,可以完全地使腫瘤移轉被觀察到;並且,相反地,在腫瘤基因已惡性轉化的細胞(oncogene-transformed cells)內阻斷TGF-β訊號時,會把這些訊號降低至比閾值更低的程度,使其無法到達癌症基因誘導癌症進行。確實如此,在鱗狀細胞癌(squamous cancers)中被逼著表現的顯性-具活性Smad2,已經被證實會和有活性的Ras共同作用,使不具侵略性的腫瘤轉變成會轉移的腫瘤;但Smad突變型不與TβRI結合,並會抑制乳癌細胞的轉移。最後,在Ras經惡性轉化的細胞中,顯性-不具活性且被截短的TβRI表現之後,則是會抑制腫瘤的產生能力(tumorigenicity)以及癌症移轉。
癌症治療後的腫瘤再增殖(repopulation)與發展,是一眾所皆知的現象。它已被證實會在放射治療、化學治療以及手術摘除後發生。(在參考文獻37中有回顧)。TGF-β的濃度會對放射線有所反應而增加,也被發現與放射治療後之肺臟受傷有關,這是由膠原蛋白沉積、肺泡壁的增厚以及內皮細胞受傷所致。放射線引起的肺臟組織傷害,會因為施以抗TGF-β抗體而顯著地減少。在手術方面,已經有人提出,因為組織操作以及傷口復原而產生的生長激素過量釋放,包括TGF-β的過量釋放,會緊接著手術之後促進癌症移轉。確實如此,手術之後,循環中隨即發現了腫瘤細胞的存在,也發現腫瘤細胞數量的增加;而且在治療性切除直腸癌兩週之後,若循環中的TGF-β濃度居高不下,就表示癌症會很快地轉移至肝臟。相反地,在手術剛切除直腸腫瘤後,血清中的TGF-β濃度卻是顯著地降低的。最後,以化學-放射治療處理後期的頭頸部癌症,以及以化學治療處理非小型細胞肺癌(non-small-cell lung cancers)的情形下,循環中的TGF-β濃度已被證實和對治療的反應相關。
在本研究中,我們展示了放射治療與化學治療會提升循環中的TGF-β濃度、循環中的癌症細胞數、以及癌症的移轉。這些效應會因為系統性地施加2G7而被阻斷,因此支持TGF-β為癌症轉移進展之肇因的說法。這些數據有數個臨床上的含意。第一,因癌症治療而增加的循環中TGF-β濃度,應該要預前地探討與監控,因為它可能是一種標記,代表腫瘤注定要在治療之後快速地增長。第二,在循環中TGF-β濃度已被觀察到增加的病人,其身上帶有的腫瘤,可能會因我們施加TGF-β抑制劑而加強基本療程的效果,也可能可以抵銷與療程相關的毒性效應,例如放射線引起的組織傷害以及纖維化。我們預期,以我們現存的方法以及尚在臨床研發中的治療用TGF-β抑制劑,這些假說在不久的將來可以被證實。
方法
藥劑與細胞株。2G7融合瘤是B. Fendly (Genentech)所贈。它是在Vanderbilt University Molecular Recognition Core facility被製出以及用親和純化法所得到的。重組的人類TGF-β1以及為TGF-β1用的Quantikine Elisa kit是從 R&D Systems取得。表現PyVmT的細胞株是從MMTV/PyVmT基因轉殖鼠的乳腺腫瘤中取得。PyVmT/TGFBR2flox/flox 及PyVmT/TGFBR2KO細胞株是分別從PyVmT/TGFBR2flox/flox 及PyVmT/TGFBR2KO老鼠的乳腺癌中取得。我們在這些老鼠中使用Cre/Lox技術,有條件地從乳腺中刪除了TGF-βII。細胞被培養在一個37°C、濕化5%的溫箱中,並且培養於添加了10% FBS(HyClone)的DMEM(Cellgro; Mediatech Inc.)中。
TGF-β1的生物檢測。貂肺臟內皮報告細胞(mink lung epithelial reporter cell)在PAI-1啟動子控制下,穩定地表現螢火蟲luciferase。這些細胞接種於12-well 培養皿上 (2x105/well)並且留置過夜供其貼附。已知濃度的可溶性TGF-β1或肺臟組織的細胞裂解液(250μg/well)加入了三次,而這些細胞被留在5%二氧化碳、37°C的環境下培養了另外24小時。在清洗之後,這些細胞被收取至200μg/well的裂解緩衝液中(Dual Luciferase Kit; Promega),luciferase的活性則是在依據廠商提供的實驗步驟下進行了測量。
DNA萃取與聚合酶連鎖反應(PCR)。以石蠟固定的腫瘤切片被取出和重新添加水分。趁著還潮濕的時候,腫瘤部分以經消毒之刀片刮取至經消毒的eppendorf管中。DNA萃取是使用Instagene (Bio-Rad)並依廠商提供的實驗步驟所完成。DNA樣本儲存在-20°C,除非要另外使用。Cre誘導的PyVmT/TGFBR2KO腫瘤重組,是用PCR引子以及前述之反應條件下所驗證的。
眼窩後(retroorbital)血液收集以及TGF-β1之定量。老鼠以1%-2% isofluorane麻醉。血液(約250μl/老鼠)採集於眼睛裡的結膜靜脈,作法是使用肝素化之Natelson tube (Fisher Scientific)然後再移轉到肝素化的(heparinized)玻璃管中。血漿的製備是使用Ficoll-Paque Plus (Amersham Biosciences)並依照廠商之使用說明進行,且增加了一個步驟,即在4°C下以10000g離心十分鐘以除去血小板。為測量PyVmT表現型細胞之培養基中的TGF-β1,1 x 106 的細胞被塗佈在含在完全培養基的10 mm盤上,並且供其貼附。該培養基在隔天被更換成無血清之培養基。隔夜培養後,細胞被施以1.25-7.5 Gy。過72小時後,培養基經收集並以快速真空法濃縮(3 ml to 500μl)。老鼠的血漿和受細胞條件培養基(cell-conditioned medium),均在下一步的TGF-β1的Quantikine Elisa kit (R&D Systems)中接受測試,依照的是廠商提供的實驗步驟。標準曲線以31.5–2,000 pg/ml的人類重組TGF-β1繪出後,被用來計算TGF-β在老鼠血漿與細胞培養基中的等量。每項標本都在另外的重製實驗中重複檢驗了三次。
TGF-β2之定量。依照廠商提供之實驗步驟,在酸活化之後,血漿標本以TGF-β2 Quantikine Elisa kit (R&D Systems)測試。標準曲線以31.5–2,000 pg/ml的人類重組TGF-β2繪出後,被用來計算TGF-β2在老鼠血漿的等量。每項標本都在另外的重製實驗中重複檢驗了三次。
老鼠的放射處理。MMTV/PyVmT基因轉殖老鼠或正常的FVB母老鼠(Harlan),在麻醉後把背部固定在鏡臺上。我們使用了單一的胸前輻射場,並使用了鉛塊掩護老鼠身體的其他部分。在某些實驗中則是使用了一個單一腹部輻射場。老鼠被施加300 kVp 的X光,其劑量為 2.05 Gy/min。所有的老鼠都依照此法規Institutional Animal Care and Use Committee of Vanderbilt University Medical Center,被留在一個專門的無病原空間中。在動物暴露在放射線或其他處理後,餵食正常食物,並且詳細觀察是否有任何情緒不穩的跡象。
小紅莓(doxorubicin)的處理。53天大的MMTV/PyVmT老鼠,經腹腔注射三次溶解於食鹽水液中的doxorubicin (Adriamycin; 5 mg/kg; Sigma-Aldrich),每次間隔21天,直到第95天止。經無效空白試劑或小紅莓處理之老鼠,其血液樣本和肺臟組織,都於第107天解剖取得。為了要檢查小紅莓處理之後的循環中腫瘤細胞,經無效空白試劑或小紅莓處理之老鼠,其血液在取得24小時後,又加入了單一劑量(5 mg/kg)。
偵測循環中腫瘤細胞。循環中腫瘤細胞之培養方法如前述,僅有少部分之修改。扼要地說,血液取得於心臟穿刺。細胞懸浮液(包括buffy coat以及紅血球)從血漿中分離出後,接種在6-well盤上,這些盤上佈有了生長激素減量之Matrigel (BD Biosciences),並添加有DMEM、10%FBS,之後便於37°C、5%二氧化碳之溫箱中培養。隔天,在每個well使用了紅血球移除緩衝液(4.15 g NH4Cl, 0.5 g NaHCO3, 0.0186 g disodium EDTA in 200 ml water)以及PBS,數次緩和的沖洗。DMEM與10%FBS(3ml/well)添加後,新鮮的培養基每三天將會補充一次。過了10-12天後,以人工計數大小為50μm以上之細胞集落。PyVmT表現是用免疫細胞化學方法來偵測的。扼要地說,吸出培養基、以PBS清洗wells,然後細胞集落以10%中性緩衝的福馬林(formalin),在室溫下固定30分鐘,再用PBS清洗2次,再於室溫下與PBS中之3%BSA共存1小時。培養盤接著培養隔夜,於4°C置放於含有生物素標記的抗PyVmT的老鼠單株抗體(diluted 1:500, BIOT-115L; Covance)。然後,加入與streptavidin結合之螢光二級抗體(Oregon Green 488; Invitrogen) 室溫下1小時,之後使用Hoechst 細胞核染色法(1 μg/ml 十使用分鐘)。免疫螢光是以Leica DM IRB 倒置顯微鏡所觀察的。
生產反轉錄病毒載體以及細胞轉導。MMTV/PyVmT細胞,是以luciferase之載體穩定轉染的。一個luciferase cassette從pGL3-Basic (Promega)裁切下來,並嵌入pMSCV-puro (Clontech)之多重複製位上。amphotropic packaging Phoenix cells 轉染後,產生感染性病毒顆粒,如前述。培養生長中(subconfluent)之癌症基因表現細胞,在4 μg/ml Polybrene (Sigma-Aldrich)中,以病毒上清液轉導6小時,而含有病毒顆粒的培養基在6小時之後更換;在48小時後,再添加含有10%已經加熱變性之FCS的DMEM,並於7-10天後將抗藥的細胞集落合併。
腫瘤產生能力以及腫瘤轉移之研究。在1%–2% isofluorane 之麻醉下,MMTV/PyVmT/Luc 細胞(2 × 106 cells in 200 μl PBS) 被注射於FVB母鼠的左側四號乳腺脂肪墊。扼要地說,在注入腫瘤細胞之前,脂肪墊便已經以手術使其暴露在外,而傷口以乾淨夾子密合,並於10天之後取走。每週以生物發光法檢查腫瘤兩次,並系列地以測徑器測量之。這些腫瘤的以立方公釐(mm3)計算之體積,是以此數學式計算:v = (w2 × l)/2,v代表體積,w代表寬度,l代表長度。當腫瘤達到了200 mm3或更大的體積(約兩週時間),老鼠就被施以放射線處理,或是不作任何處理。老鼠繼續每週兩次接受生物發光法的監控觀察,直到實驗終結。兩週之後,將老鼠犧牲並取得基本的腫瘤與肺臟。以解剖顯微鏡計算肺臟表面之癌症轉移數,之後這些肺臟以10%中性緩衝之福馬林(formalin)固定之。固定好的腫瘤切片,以H&E染色之,並以顯微鏡檢查是否有肺癌之轉移。血液是由心臟穿刺取得以如前述,測量TGF-β之濃度以及循環中之腫瘤細胞數。
在其他的情形裡,我們使用了MMTV/PyVmT、PyVmT/TGFBR2flox/flox或PyVmT/TGFBR2KO細胞來表現luciferase。2.5 × 106 之單一細胞懸浮液製備後,重新溶於無菌之PBS液中。0.5 × 106 (in 200 μl)的細胞被注射入FVB母鼠之側面尾部靜脈中。靜脈注射24小時後,腫瘤細胞於老鼠肺部之位置,以生物發光法確定之,並且自此之後每週追蹤兩次。為了要在活體內阻斷TGF-β,在把10 Gy注入老鼠胸腔的前24小時,將15 mg/kg的2G7注入老鼠腹腔內,每週兩次,持續進行,直到老鼠死亡。
生物發光法顯影技術。要將生物發光之訊號與腫瘤負載量建立定量的對應關係,先以溶解於去離子水的 luciferin substrate (d-luciferin potassium salt; Promega)進行腹腔注射(0.15 mg/g body weight),在注射之後12分鐘內取得影像。根據我們之前的時間記錄之實驗,在此時間記錄訊號高峰處之強度(資料未呈現)。在取得影像時,將老鼠以isofluorane麻醉。生物發光顯影是以Vanderbilt University Small Animal Imaging Center的 IVIS-200 顯影系統 (Xenogen)完成的。我們採用了3-5分鐘的整合計算時間,並使用on-chip binning of 8,以增加訊號-雜訊比。生物發光顯影之定量分析是以Living Image software (Xenogen)完成,我們在此定義了感興趣區域(regions of interest,ROI)並測量了integrated photon counts。
統計。所有的實驗數據均以雙尾Student’s t test分析。P值小於0.05被視為顯著。
表一
穩定表現luciferase的MMTV/PyVmT細胞,被注射入FVB老鼠的右側鼠蹊乳腺脂肪墊中。擁有200mm3以上腫瘤的老鼠,不加以放射處理,或是在胸腔施加10
Gy的放射處理。在24小時及2週後,以心臟穿刺取得血液,並測量細胞懸浮液在活體外形成細胞集落的能力,如「方法」文中所示。數據為兩個分別的實驗中,每組五隻老鼠的平均值±標準差。RT是指放射治療(radiation
therapy)。在同一時間點下,AP
= 0.016 versus 0 Gy。
圖一
放射線與化學治療增加循環中TGF-β1濃度。 (A) 10 Gy 的放射線被施加於FVB老鼠之胸腔(thorax) (左)或骨盆腔(pelvis )(右)。24小時之後取得血液,血漿中之TGF-β1濃度以在「方法」中所述測量。(B) 八週大、帶有腫瘤之MMTV/PyVmT 老鼠,或非基因轉殖之FVB 老鼠,在四號乳腺脂肪墊帶有200mm3以上大小之腫瘤,不作任何處理或施加10 Gy 於胸腔。血漿中TGF-β1 濃度,24小時後測量之。(C) 在第八週起,每隔21天,以空白試劑或腹腔注射小紅莓 (5 mg/kg i.p.),處理基因轉殖老鼠三次。血漿中TGF-β1濃度在第十五週測量之。A–C 之數據代表三個不同的獨立實驗,每組使用三個實驗對象。(D) FVB老鼠胸腔被施加10 Gy。五週後,取得經放射線處理、控制組老鼠之肺臟,及其細胞裂解液(250 μg/ml),加到含有貂肺上皮細胞的triplicate wells,這些細胞會穩定地表現plasminogen activator inhibitor–1/luciferase reporter (PAI-1/luciferase reporter)。二十四小時後,測量luciferase之表現,方法如「方法」部分中所述。圖示:*表示P < 0.05, **表示P < 0.01, ***表示P < 0.001 相對於控制組。
圖二
放射線與化學治療增加循環中腫瘤細胞量以及肺部之腫瘤轉移。(A) MMTV/PyVmT母鼠在八週大時,接受胸腔放射線處理。在第十三週時,當實驗終結,以心臟穿刺取得血液,並量取其細胞懸浮液在體外產生細胞集落之能力,如「方法」中所述。CTCs代表循環中腫瘤細胞(circulating tumor cells)。在下面為一些具有代表性的影像,是由血液中取得、循環中單一個腫瘤細胞形成的細胞集落。基因轉殖成功的細胞集落,用PyVmT抗體和螢光之次級抗體處理,並以螢光顯微鏡觀察並估計之。(B) 與A 同樣之老鼠,在第十三週大時計算肺部表面癌症之轉移數目。A和B中之數據是代表三個獨立的實驗,每組中有四隻老鼠。從控制組以及經放射線處理之基因轉殖老鼠上,取得具代表性之 H&E 染色的肺部組織切片。這些切片是胸腔放射處理後五週所取得,如下方所示。黑色箭頭所指為肺部癌症轉移。(C) 八週大、帶有腫瘤的 MMTV/PyVmT 基因轉殖老鼠,每隔21天以空白試劑或腹腔注射小紅莓(5 mg/kg i.p.)處理共三次。此實驗於第十五週時終結,並計算肺部表面之癌症轉移數量。具代表性之H&E染色肺部切片,其含有癌症轉移部分的中心如右方圖示。原始放大倍率為100倍。*表示P < 0.05, **表示P < 0.01 ,相較於控制組。
圖三
胸腔放射線會增加殖入腫瘤至肺部的癌症轉移。(A) 穩定表現luciferase的MMTV/PyVmT 細胞,被注入同源FVB老鼠之乳腺脂肪墊中。之後,兩週一次,以生物發光顯影法監測腫瘤生長以及肺部癌症轉移。圖示為一具代表性之老鼠,圖為細胞注入兩週之後所攝(B與C)。帶有至少200 mm3 PyVmT/Luc腫瘤的老鼠,以胸腔放射線(10 Gy)處理之或不施加任何處理。 (B) 肺部癌症轉移在兩週之後定量。數據為每兩次實驗中之一次,每組五隻老鼠之平均值 ± 標準差。**表示P < 0.01 相較於控制組 (C) 肺部與原發腫瘤之H&E 切片。原始放大倍率為100倍。
圖四
放射線的先行處理,會使移轉的肺癌細胞更能夠拓展在原本無腫瘤的老鼠身上。(A) MMTV/PyVmT 細胞,在加有不含血清之培養基之100-mm 培養皿裡,以1.25–7.5 Gy處理。受細胞所影響之培養基在72小時後進行收集,並以如「方法」中所述之ELISA得知TGF-β1之濃度。(B) 穩定表現luciferase之MMTV/PyVmT 細胞,由尾部靜脈注入未交配之FVB母鼠。在接受細胞注射1小時之前,老鼠在圖中所標部分之胸部,以10 Gy處理之。在注射之後2週,肺部之癌症細胞以老鼠的生物發光法進行視覺觀察(頂圖)。某些情形中,肺臟在添加d-luciferin之後被手術移除,並於體外進行顯影(底圖)。控制組如左圖所示,受放射線處理之老鼠如右圖所示。(C) 具代表性的整體肺部載片(頂圖)以及控制組與放射線處理組之H&E肺部切片(底圖;原放大倍率為100)。(D) 在控制組與放射線處理老鼠中,肺部表面癌症轉移之定量(左圖)與肺部重量(右圖)。數據為兩個獨立實驗中,每組的五隻老鼠的平均值±標準差。圖示:**表示P < 0.01, ***表示P < 0.001 ,相較於控制組。
圖五
TGF-β中和抗體2G7,阻斷了放射線誘導增加的肺部癌症轉移。(A和B) 八週大、 帶有腫瘤的MMTV/PyVmT老鼠於胸腔接受10 Gy處理。在圖中標出之部分,老鼠被每週兩次施以15 mg/kg 的2G7,直到第十三週。此時進行肺部表面之癌症轉移計數。(A) 數據為每組中五隻老鼠的平均值 ± 標準差。(B) 代表性的H&E 染色肺部切片。此實驗重複一次取得了類似的結果。(C) 當實驗終結,以心臟穿刺取得血液,並量取其細胞懸浮液在體外產生細胞集落之能力,如「方法」部分中所述。(D) 在第十三週,經過胸腔放射處理、帶有腫瘤的基因轉殖鼠,收集血液。細胞懸浮液如C圖中情形,在20 μg/ml 2G7或PBS環境下,塗佈於培養盤上。10-12天後,人工計數等於或大於50 μm的細胞集落。數據為每組五隻老鼠的平均值±標準差。*表示P < 0.05, **表示P < 0.01 相較於控制組。
圖六
腫瘤細胞中不存在TGFβRII,能抵銷受放射線誘發的肺部癌症轉移之增加。(A–C) 未交配的、八週大的FVB母鼠,從尾部靜脈注入穩定表現 luciferase的PyVmT/TGFBR2flox/flox 與 PyVmT/TGFBR2KO 細胞。這些老鼠在注射腫瘤細胞之前,一組接受、一組未接受胸腔的10 Gy處理。兩週後,以生物發光(A)、組織染色(B)、以及人工計數肺部表面癌症轉移(C)的方法,來測定肺部表面癌症轉移,如「方法」中所述。資料為每組中四隻老鼠的平均值± 標準差。 (D) 兩組細胞株的基因體DNA之PCR結果,顯示重組片段的僅存在於PyVmT/TGFBR2KO 細胞中。**表示P < 0.01 相較於控制組。
譯者:Jeffrey Chu at Hsinchu, Taiwan. 2009/4/4
不免有所遺漏、錯誤,敬請參考與指正,謝謝。